Back in the lab
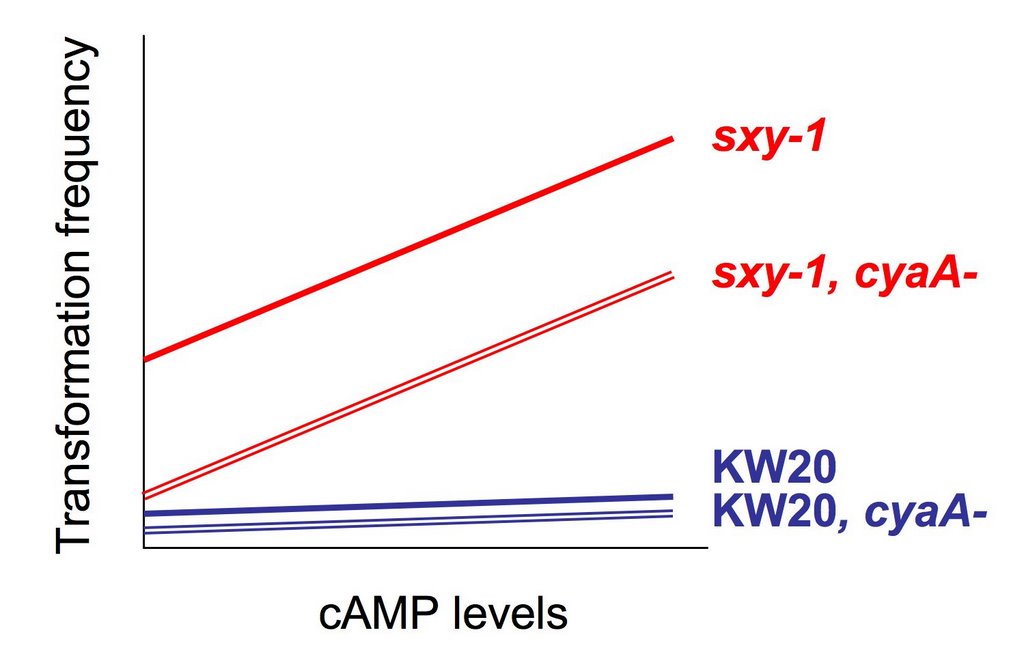
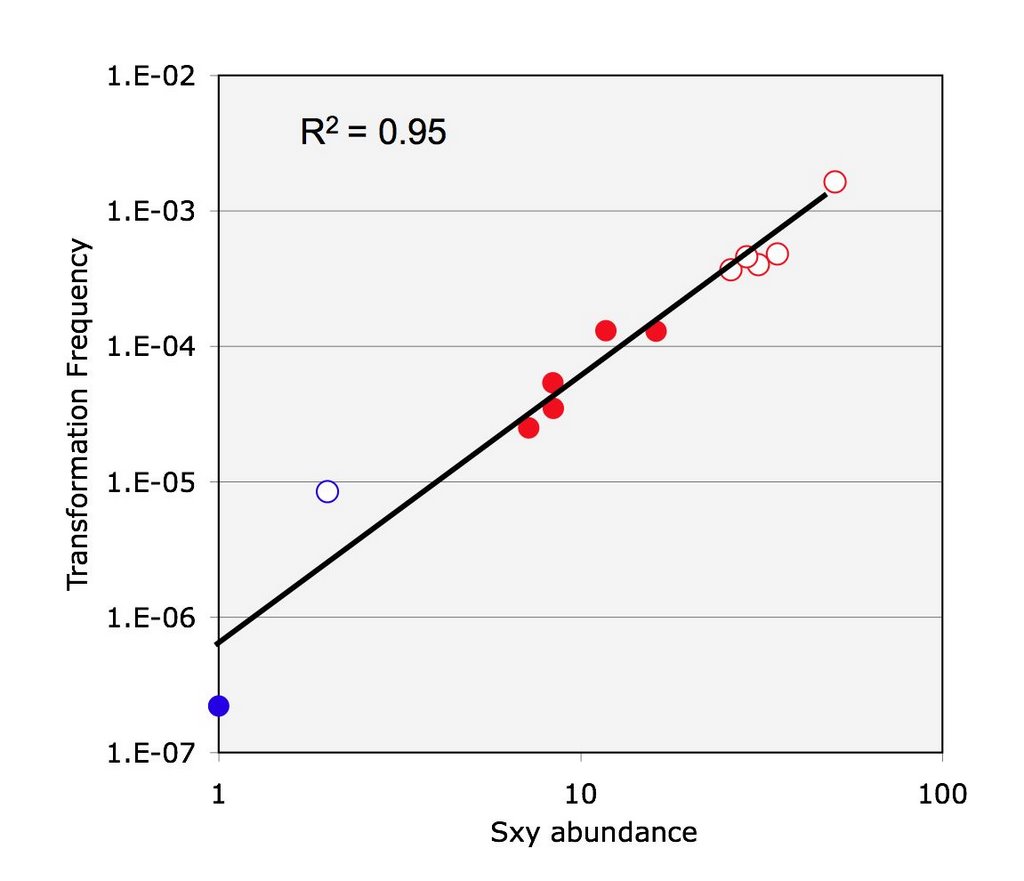
I am currently planning my experiments for the next couple of months so that I know exactly what I want to be doing each day. My first goal is to resolve a problem that arose just before my departure to Inuvik. I was repeating an experiment to quantify the relative levels of sxy mRNA in strains that have regulatory mutations in the sxy gene.
There are several steps required to do this. First, cells are cultured and all RNA is extracted. The RNA samples are treated with DNase, then random hexamer-primed reverse transcription is used to convert the RNA into cDNA. The cDNA is then used as template in a SYBR green real time PCR assay. In this assay, more starting template results in the synthesis of more amplicon early in the PCR cycles. Amplicons are bound by the SYBR green fluorophore and fluorescence increases; fluorescence is measured each thermocycle. There are many steps where problems can arise, but appropriate checks and controls are used throughout. First, a gel is run to test the quality of the extracted RNA. Later in the final PCR reactions, non-DNase treated RNA is included as a template to ensure 1) that there is more template in reverse transcription reactions, confirming that the RT step worked, and 2) that chromosomal DNA is not a significant contaminant. Also, chromosomal DNA is used to confirm that the PCR Master mix can amplify target genes.
In the past we have had problems with chromosomal DNA contamination during the RNA extraction/purification, but that has long since been remedied. On Friday Oct. 13, I encountered a whole new problem: no amplification was detected in any of the real time PCR reactions. One odd result is that the fluorescence levels were very high throughout the run, suggesting that there may have been amplification as usual, but that the Master Mix fluorophores or the detector are not working properly. There are other possible suspects: 1. The least probable problem is that the primers have gone bad (I know they were added to the Master Mix), but they worked beautifully a couple of months ago. 2. I used a new batch of SYBR Master Mix, and it may have bad Taq enzyme or bad fluorohpores. 3. The machine is not thermocycling or its detector is not working properly.
My first troubleshooting step is to run some of the PCR reactions on a gel to see if amplification did occur. If yes, fluorescence is the problem: either the detector or the MMix is not working. If no, Taq or the thermocycle is not working. Either way, the second step will be to run a couple of reactions comparing the new Master Mix to older leftover MMix. Also, I will include another primer set to test whether my primers are the problem. If I again see no amplification, I will try the reactions again on a different realtime PCR machine.
On the positive side, my cDNA samples are already prepared. Once I solve the PCR problem, it will be relatively quick to measure sxy mRNA levels as I had originally set out to do.
posted by Some call me Tim @ 1:23 PM
2 comments
![]()